MIO-A
Human movements are still poorly understood, mainly due to their high complexity and large number of degrees of freedom embedded in neural commands. This shortcoming can be compensated by analysing electrical activity of muscles at the surface of the skin, so called surface electromyogram (EMG). The latter offers numerous advantages over clinically more established intramuscular investigations, such as repeatability of measurements, patient’s comfort, no risk of infection and lower examination costs.
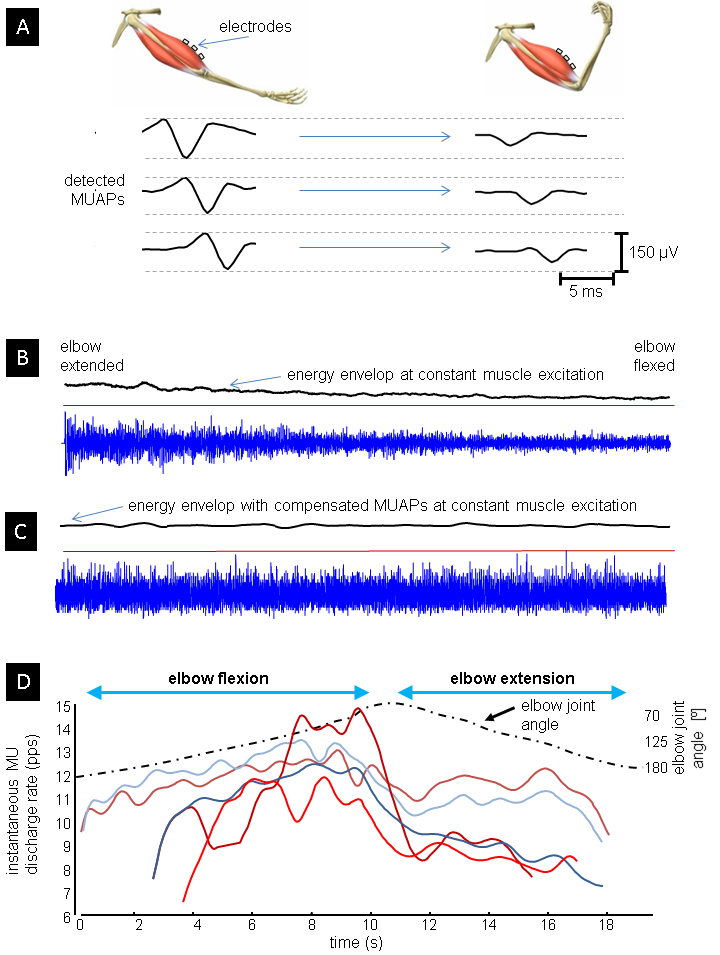
Surface EMG has been extensively used in the fields of neuroscience, rehabilitation, pathophysiological investigations, training of the athletes and in man-machine interfacing. The main challenge in all these applications is identification of neural commands out of recorded EMG. Namely, EMG is composed of many action potentials (AP) being contributed by basic functional units of a muscle, so called motor units (MUs). Central nervous system controls muscle force by controlling the number of active MUs and their firing rates, whereas the shape of MU APs is completely irrelevant from the central control viewpoint.
Moreover, surface EMG appears to be highly interferential and the muscle activations are typically estimated by calculating the EMG’s energy envelops. This approach is sensitive to the shape of MU APs, which depend on muscle anatomy and geometry and are one of the main reasons for reported large variability and low repeatability of EMGs. This negative impact of MU APs has been largely ignored in the studies of motor behaviour. Even more, EMG envelopes have been used to establish a theory of muscle synergies, which hypothesizes that a shared co-activation of several muscles is modulated by a single neural command.
We analysed the methodological deficiencies of the currently established theory of muscular synergies and showed that established methodologies are sensitive to geometric changes in muscles, especially when changes in muscular excitation are small. We have demonstrated that the current methodologies yield statistically significant false estimates of muscular co-activation in robotic-supported rehabilitation after stroke and that the procedures developed in this project significantly improve this estimates.
In order to demonstrate their robustness and suitability for analysis of central nervous system disorders, the developed techniques will be used to accurately track the pathological variability in excitation of individual upper limb muscles in the hamiparetic patients and to compare this variability to the functional movement errors as assessed by a haptically-controlled UHD rehabilitation robot. This is expected to provide better information support to rehabilitation decisions and, thus, maximize the patient’s rehabilitation potential.
The developed methodology is also time effective and supports a variety of measuring electrodes per muscle. It greatly improves the accuracy of assessing muscle activity on the surface of the skin and has great potential in the fields of rehabilitation, athletes' training, ergonomics, prosthetics, orthotics and the general field of muscle-machine interfaces.